Energia para a vida
Um dos principais fatores limitantes a vida dos seres vivos é a obtenção de energia para as suas atividades. De acordo com a teoria heterotrófica, os primeiros seres vivos seriam procariontes heterotróficos vivendo num meio aquático, de onde retirariam nutrientes, formados na atmosfera e acumulados nos lagos e oceanos primitivos. Devido á sua grande simplicidade, estes seres utilizariam processos igualmente rudimentares para retirar energia dessas moléculas de que se alimentavam. Esse mecanismo seria semelhante à fermentação realizada ainda por muitos organismos atuais.
Há mais de 2 bilhões de anos, surgiram os primeiros organismos autotróficos, procariontes capazes de produzir o seu próprio alimento através da fotossíntese. Este processo revolucionário, além de permitir a sobrevivência dos autotróficos, também serviu aos heterotróficos, que passaram a alimentar-se deles.
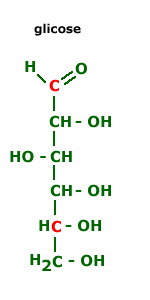
Os organismos retiram energia das mais diversas moléculas orgânicas (açucares, aminoácidos, ácidos graxos, etc.), mas a glicose é a mais freqüente, tanto na fermentação como na respiração. Para a fermentação ou respiração os organismos heterotróficos obtém a glicose se alimentado dos únicos que produzem glicose, os organismos autotróficos fotossintetizantes.  Atualmente, apenas algumas bactérias e fungos utilizam o processo de fermentação para obter energia. Todos os outros organismos, sejam autótrofos (algas e plantas) ou heterótrofos (algumas bactérias, fungos e protozoários e animais), se utilizam da respiração aeróbica, um processo de obtenção de energia muito mais eficiente do que a fermentação.
A glicose e o metabolismo
Associe anabolismo a síntese e catabolismo a decomposição de substâncias. De modo geral essas duas modalidades ocorrem juntas.
Durante o catabolismo, que ocorre nos processos energéticos, por exemplo, a energia liberada em decorrência da utilização dos combustíveis biológicos poderá ser canalizada para as reações de síntese de outras substâncias, que ocorre no anabolismo.
Energia sob a forma de ATP
Cada vez que ocorre a desmontagem da molécula de glicose, a energia não é simplesmente liberada para o meio. A energia é transferida para outras moléculas (chamadas de ATP - Adenosina Trifosfato), que servirão de reservatórios temporários de energia, “bateriazinhas” que poderão liberar “pílulas” de energia nos locais onde estiverem.
No citoplasma das células é comum a existência de uma substância solúvel conhecida como adenosina difosfato, ADP. É comum também a existência de radicais solúveis livres de fosfato inorgânico (que vamos simbolizar por Pi), ânions monovalentes do ácido orto-fosfórico. Cada vez que ocorre a liberção de energia na respiração aeróbica, essa energia liga o fosfato inorgânico (Pi) ao ADP, gerando ATP. Como o ATP também é solúvel ele se difunde por toda a célula. 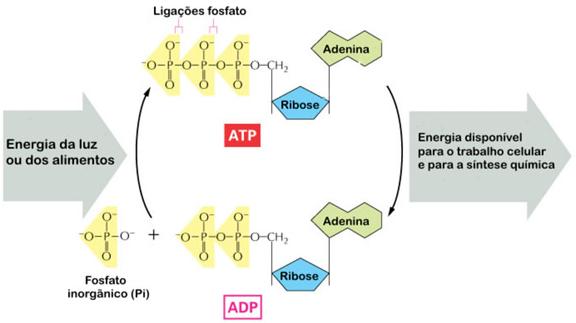
A ligação do ADP com o fosfato é reversível. Então, toda vez que é necessário energia para a realização de qualquer trabalho na célula, ocorre a conversão de algumas moléculas de ATP em ADP + Pi e a energia liberada é utilizada pela célula. A recarga dos ADP ocorre toda vez que há liberação de energia na desmontagem da glicose, o que ocorre na respiração aeróbia ou na fermentação.
A estrutura do ATP
O ATP é um composto derivado de nucleotídeo em que a adenina é a base e o açúcar é a ribose. O conjunto adenina mais ribose é chamado de adenosina. A união de adenosina com três radicais fosfato leva ao composto adenosina trifosfato, ATP. As ligações que mantêm o segundo e o terceiro radicais fosfato presos no ATP são altamente energéticas (liberam cerca de 7 Kcal/mol de substância). 
Fermentação
Liberação de energia através da fermentação
A fermentação é um processo de liberação de energia que ocorre sem a participação do oxigênio(processo anaeróbio). A fermentação compreende um conjunto de reações enzimaticamente controladas, através das quais uma molécula orgânica é degradada em compostos mais simples, liberando energia. A glicose é uma das substâncias mais empregadas pelos microorganismos como ponto de partida na fermentação.
Glicólise
Na glicólise, cada molécula de glicose é desdobrada em duas moléculas de piruvato (ácido pirúvico), com liberação de hidrogênio e energia, por meio de várias reações químicas.
O hidrogênio combina-se com moléculas transportadores de hidrogênio (NAD), formando NADH + H+, ou seja NADH2. 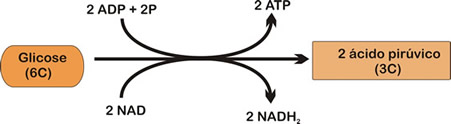
Tipos de Fermentação
Fermentação Alcoólica
As leveduras e algumas bactérias fermentam açucares, produzindo álcool etílico e gás carbônico (CO2), processo denominado fermentação alcoólica.
Na fermentação alcoólica, as duas moléculas de ácido pirúvico produzidas são convertidas em álcool etílico (também chamado de etanol), com a liberação de duas moléculas de CO2 e a formação de duas moléculas de ATP. 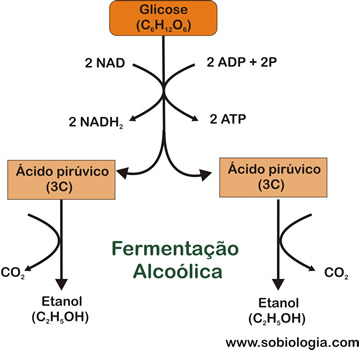
Esse tipo de fermentação é realizado por diversos microorganismos, destacando-se os chamados “fungos de cerveja”, da espécie Saccharomyces cerevisiae. O homem utiliza os dois produtos dessa fermentação: o álcool etílico empregado há milênios na fabricação de bebidas alcoólicas (vinhos, cervejas, cachaças etc.), e o gás carbônico importante na fabricação do pão, um dos mais tradicionais alimentos da humanidade. Mais recentemente tem-se utilizado esses fungos para a produção industrial de álcool combustível.
|
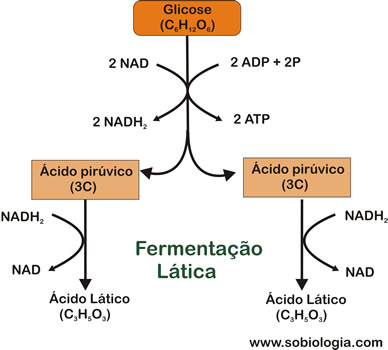
Fermentação Lática
Os lactobacilos (bactérias presentes no leite) executam fermentação lática, em que o produto final é o ácido lático. Para isso, eles utilizam como ponto de partida, a lactose, o açúcar do leite, que é desdobrado, por ação enzimática que ocorre fora das células bacterianas, em glicose e galactose. Aseguir, os monossacarídeos entram nas células, onde ocorre a fermentação.
Cada molécula do ácido pirúvico é convertido em ácido lático, que também contém três átomos de carbono.
O sabor azedo do leite fermentado se deve ao ácido lático formado e eliminado pelos lactobacilos. Oabaixamento do pH causado pelo ácido lático provoca a coagulação das proteínas do leite e a formação do coalho, usado na fabricação de iogurtes e queijos.
Fermentação láctica no homem!
 |
Você já deve ter ouvido que é comum a produção de ácido lático nos músculos de uma pessoa, em ocasiões que há esforço muscular exagerado. A quantidade de oxigênio que as células musculares recebem para a respiração aeróbia é insuficiente para a liberação da energia necessária para a atividade muscular intensa.
Nessas condições, ao mesmo tempo em que as células musculares continuam respirando, elas começam a fermentar uma parte da glicose, na tentativa de liberar energia extra.
O ácido láctico acumula-se no interior da fibra muscular produzindo dores, cansaço e cãibras.
Depois, uma parte desse ácido é conduzida pela corrente sanguínea ao fígado onde é convertido em ácido pirúvico.
|
Fermentação Acética
As acetobactérias fazem fermentação acética, em que o produto final é o ácido acético. Elas provocam o azedamento do vinho e dos sucos de frutas, sendo responsáveis pela produção de vinagres.
Respiração aeróbica
Os processos fermentativos levam a formação de moléculas orgânicas pequenas, mas ainda capazes de liberar energia. Por exemplo, o álcool etílico, um dos produtos da fermentação da glicose, contêm quantidades razoáveis de energia liberáveis, tanto que é utilizado como combustível.
A respiração aeróbia consiste em levar a diante o processo de degradação das moléculas orgânicas, reduzindo-as as moléculas praticamente sem energia liberável. Os produtos da degradação inicial da molécula orgânica são combinados com o oxigênio do ar e transformados em gás carbônico e água.
|
O rendimento energético da respiração aeróbica
O processo de respiração aeróbica, é muito mais eficiente que a da fermentação: para cada molécula de glicose degradada, são produzida na respiração, 38 moléculas de ATP, a partir de 38 moléculas de ADP e 38 grupos de fosfatos. Na fermentação, apenas duas moléculas de ATP são produzidas para cada molécula de glicose utilizada. A eficiência da respiração em termos energéticos é, portanto, dezenove vezes maior do que a da fermentação.
A respiração aeróbica é um processo muito mais complexo que a fermentação. São necessários cerca de 60 passos metabólicos a mais, além dos nove que compõe a glicólise, para que uma molécula de glicose seja totalmente degradada a CO2 e H2O, em presença de O2.
Etapas da respiração aeróbica
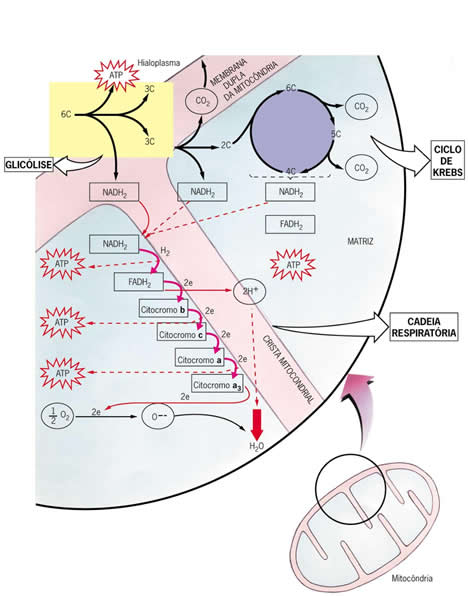
A degradação da glicose na respiração celular se dá em três etapas fundamentais: glicólise, ciclo de Krebs e cadeia respiração. A glicólise ocorre no hialoplasma da célula, enquanto o ciclo de Krebs e a cadeia respiratória ocorrem no interior das mitocôndrias.
Glicólise
Como já vimos, a glicólise consiste na transformação de uma molécula de glicose, ao longo de várias etapas, em duas moléculas de ácido pirúvico.
Nesse processo são liberados quatro hidrogênios, que se combinam dois a dois, com moléculas de uma substância celular capaz de recebê-los: o NAD (nicotinamida-adenina-dinucleotídio). Ao receber os hidrogênios, cada molécula de NAD se transforma em NADH2. Durante o processo, é liberada energia suficiente para a síntese de 2 ATP.
Nesse processo são liberados quatro hidrogênios, que se combinam dois a dois, com moléculas de uma substância celular capaz de recebê-los: o NAD (nicotinamida-adenina-dinucleotídio). Ao receber os hidrogênios, cada molécula de NAD se transforma em NADH2. Durante o processo, é liberada energia suficiente para a síntese de 2 ATP.
Ciclo do Ácido Cítrico ou de Krebs
Oxidação do Ácido Pirúvico
As moléculas de ácido pirúvico resultantes da degradação da glicose penetram no interior das mitocôndrias, onde ocorrerá a respiração propriamente dita. Cada ácido pirúvico reage com uma molécula da substância conhecida como coenzima A, originando três tipos de produtos: acetil-coenzima A, gás carbônico e hidrogênios.
O CO2 é liberado e os hidrogênios são capturados por uma molécula de NADH2 formadas nessa reação. Estas participarão, como veremos mais tarde, da cadeia respiratória.
Em seguida, cada molécula de acetil-CoA reage com uma molécula de ácido oxalacético, resultando em citrato (ácido cítrico) e coenzima A, conforme mostra a equação abaixo:
Em seguida, cada molécula de acetil-CoA reage com uma molécula de ácido oxalacético, resultando em citrato (ácido cítrico) e coenzima A, conforme mostra a equação abaixo:
1 acetil-CoA + 1 ácido oxalacético  1 ácido cítrico + 1 CoA
1 ácido cítrico + 1 CoA
(2 carbonos) (4 carbonos) (6 carbonos)
(2 carbonos) (4 carbonos) (6 carbonos)
Analisando a participação da coenzima A na reação acima, vemos que ela reaparece intacta no final. Tudo se passa, portanto, como se a CoA tivesse contribuído para anexar um grupo acetil ao ácido oxalacético, sintetizando o ácido cítrico.
Cada ácido cítrico passará, em seguida, por uma via metabólica cíclica, denominada ciclo do ácido cítrico ou ciclo de Krebs, durante o qual se transforma sucessivamente em outros compostos.
Cada ácido cítrico passará, em seguida, por uma via metabólica cíclica, denominada ciclo do ácido cítrico ou ciclo de Krebs, durante o qual se transforma sucessivamente em outros compostos.
Analisando em conjunto as reações do ciclo de Krebs, percebemos que tudo se passa como se as porções correspondentes ao grupo acetil, anteriormente transferidas pela CoA, fossem expelidas de cada citrato, na forma de duas moléculas de CO2 e quatro hidrogênios. Um citrato, sem os átomos expelidos, transforma-se novamente em ácido oxalacético.
Os oito hidrogênios liberados no ciclo de Krebs reagem com duas substâncias aceptoras de hidrogênio, o NAD e o FAD, que os conduzirão até as cadeias respiratórias, onde fornecerão energia para a síntese de ATP. No próprio ciclo ocorre, para cada acetil que reage, a formação de uma molécula de ATP.
Cadeia respiratória e liberação de energia
O destino dos hidrogênios liberados na glicólise e no ciclo de Krebs é um ponto crucial no processo de obtenção de energia na respiração aeróbica.
Como vimos, foram liberados quatro hidrogênios durante a glicólise, que foram capturados por duas moléculas de NADH2. Na reação de cada ácido pirúvico com a coenzima A formam-se mais duas moléculas de NADH2. No ciclo de Krebs, dos oito hidrogênios liberados, seis se combinam com três moléculas de NAD, formando três moléculas de NADH2, e dois se combinam com um outro aceptor, o FAD, formando uma molécula de FADH2.
Através de sofisticados métodos de rastreamento de substâncias, os bioquímicos demonstraram que os hidrogênios liberados na degradação das moléculas orgânicas e capturados pelos aceptores acabam por se combinar com átomos de oxigênio provenientes do O2 atmosférico. Dessa combinação resultam moléculas de água.
Antes de reagirem como o O2, porém, os hidrogênios, percorrem uma longa e complexa trajetória, na qual se combinam sucessivamente com diversas substâncias aceptoras intermediárias. Ao final dessa trajetória, os hidrogênios se encontram seus parceiros definitivos, os átomos de oxigênio do O2. Esse conjunto de substâncias transportadoras de hidrogênio constitui a cadeia respiratória.
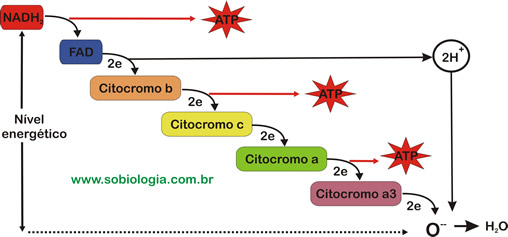
Se os hidrogênios liberados na degradação das moléculas orgânicas se combinassem direta e imediatamente com o O2, haveria desprendidamente de enorme quantidade de energia em forma de calor, impossível de ser utilizada. Para contornar esse problema, as células utilizam um mecanismo bioquímico que permite a liberação gradual de energia. Tudo se passa como os hidrogênios descessem uma escada, perdendo energia a cada degrau. Liberada em pequenas quantidades, a energia pode ser, então, utilizada na síntese de moléculas de ATP, a partir de ADP e fosfatos.
Aceptores de hidrogênio da cadeia respiratória
As moléculas de NAD, de FAD e de citocromos que participam da cadeia respiratória captam hidrogênios e os transferem, através de reações que liberam energia, para um aceptor seguinte. Os aceptores de hidrogênio que fazem parte da cadeia respiratória estão dispostos em sequência na parede interna da mitocôndria.
O ultimo aceptor de hidrogênios na cadeia respiratória é a formação de moléculas de ATP, processo chamado de fosforilação oxidativa. Cada molécula de NADH2 que inicia a cadeia respiratória leva à formação de três moléculas de ATP a partir de três moléculas de ADP e três grupos fosfatos como pode ser visto na equação a seguir:
1 NADH2 + ½ O2 + 3 ADP + 3P 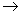 1 H2O + 3 ATP + 1 NAD
1 H2O + 3 ATP + 1 NAD
Já a FADH2 formado no ciclo de Krebs leva à formação de apenas 2 ATP.
1 FADH2 + ½ O2 + 2 ADP + 2P 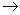 1 H2O + 2 ATP + 1 FAD
1 H2O + 2 ATP + 1 FAD
Contabilidade energética da respiração aeróbica
Na glicólise há um rendimento direto de duas moléculas de ATP por moléculas de glicose degradada. Formam-se, também, duas moléculas de NADH2 que, na cadeia respiratória, fornecem energia para a síntese de de seis moléculas de ATP.
Durante o ciclo de Krebs, as duas moléculas de Acetil-CoA levam a produção direta de duas moléculas de ATP. Formam-se, também, também, seis moléculas de NADH2 e duas moléculas de FADH2 que, na cadeia respiratória, fornecem energia para a síntese de dezoito moléculas de ATP (para o NAD) equatro moléculas de ATP (para o FAD).
A contabilidade energética completa da respiração aeróbica é, portanto: 2 + 6 + 6 + 2 + 18 + 4 = 38 ATP. O resumo de todas as etapas resulta na seguinte equação geral:
1 C6H12O6 + 6 O2 + 38 ADP + 38 P 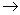 6 CO2 + 6 H2O + 38 ATP
6 CO2 + 6 H2O + 38 ATP
A importância metabólica do ciclo de Krebs
Ao estudarmos a respiração aeróbica, partimos de moléculas de glicose. Outras substâncias, porém, como proteínas e gorduras, também podem servir de combustível energético. Depois de devidamente transformadas, essas substâncias produzem moléculas de acetil, o combustível básico do ciclo de Krebs.
O ciclo de Krebs é a etapa da respiração em que a acetil-CoA oriunda das moléculas alimentares é “desmontada” em CO2 e H2O, e a energia produzida é usada na síntese de ATP.
O ciclo de Krebs é a etapa da respiração em que a acetil-CoA oriunda das moléculas alimentares é “desmontada” em CO2 e H2O, e a energia produzida é usada na síntese de ATP.
Porém o ciclo de Krebs não participa apenas do metabolismo energético: à medida que as diversas substâncias do ciclo vão se formando, parte delas pode ser “desviada”, indo servir de matéria-prima para a síntese de substâncias orgânicas (anabolismo).
Por exemplo, uma parte das substâncias usadas pelas células para produzir aminoácidos, nucleotídeos e gorduras provém do ciclo de Krebs.
Veja na figura abaixo as etapas da respiração celular e a sua localização:
Fotossíntese
A fotossíntese é o principal processo autotrófico e é realizada pelos seres clorofilados, representados porplantas, alguns protistas, bactérias fotossintetizantes e cianobactérias.
Na fotossíntese realizada pelos seres fotossintetizantes, com exceção das bactérias, gás carbônico (CO2) e água (H20) são usados para a síntese de carboidratos, geralmente a glicose. Nesse processo há a formação de oxigênio (O2), que é liberado para o meio.
|
A fotossíntese realizada pelas bactérias fotossintetizantes difere em muitos aspectos da realizada pelos demais organismos fotossintetizantes, como veremos a seguir.
A fórmula geral da produção de glicose pela fotossíntese dos eucariotos e cianobactérias é:
6 CO2 + 12 H2O 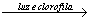 C6H12O6 + 6 O2 + H2O
C6H12O6 + 6 O2 + H2O
Essa equação mostra que, na presença de luz e clorofila, o gás carbônico e a água são convertidos em uma hexose – neste exemplo, a glicose - havendo liberação de oxigênio.
Os seres fotossintetizantes são fundamentais para a manutenção da vida em nosso planeta, pois são a base da maior parte das cadeias alimentares e produzem oxigênio, gás mantido na atmosfera em concentrações adequadas graças principalmente a atividade fotossintética.
Origem do oxigênio e fotossíntese bacteriana
O oxigênio liberado pela fotossíntese realizada pelos eucariontes e pelas cianobactérias provém da água, e não do gás carbônico, como se pensava antigamente.
O primeiro pesquisador a propor isso foi Cornelius Van Niel, na década de 1930, quando estudava bactérias fotossintetizantes. Ele verificou que as bactérias vermelhas sulfurosas (ou tiobactérias púrpuras) realizavam uma forma particular de fotossíntese em que não havia necessidade de água nem formação de oxigênio. Essas bactérias usam gás carbônico e sulfeto de hidrogênio (H2S) e produzem carboidrato e enxofre.
Van Niel escreveu, então, a fórmula geral da fotossíntese realizada por essas bactérias:
Fotossíntese bacteriana
6 CO2+ 2 H2S CH2O + H2O + 2 S
CH2O + H2O + 2 S
6 CO2+ 2 H2S
Foi a compreensão desse processo de fotossíntese que levou o pesquisador a propor a equação geral da fotossíntese:
6 CO2+ 2 H2A
Essa equação mostra que H2A pode ser a água (H2O) ou o sulfeto de hidrogênio (H2S) e evidencia que, se for água ela é a fonte de oxigênio na fotossíntese.
Essa interpretação foi confirmada posteriormente, na década de 1940, por experimentos em que pesquisadores forneciam às plantas água cujo oxigênio era de massa 18 (O18, isótopo pesado do oxigênio) em vez de 16 (O16), como o oxigênio da água comum. Eles verificaram que o oxigênio liberado pela fotossíntese era o O18, corroborando a interpretação de Van Niel.
Essa interpretação foi confirmada posteriormente, na década de 1940, por experimentos em que pesquisadores forneciam às plantas água cujo oxigênio era de massa 18 (O18, isótopo pesado do oxigênio) em vez de 16 (O16), como o oxigênio da água comum. Eles verificaram que o oxigênio liberado pela fotossíntese era o O18, corroborando a interpretação de Van Niel.
Ficou comprovado, então, que o oxigênio liberado durante a fotossíntese dos eucariontes e das cianobactérias provém da água e não do gás carbônico.
Os Cloroplastos
Os plastos ou plastídeos é um grupo de organelas específicas de células vegetais, que possuem características semelhantes com as mitocôndrias como: membrana dupla, DNA próprio e origem endosimbionte.
Os plastos desenvolvem-se a partir de proplastídeos, que são organelas pequenas presentes nas células imaturas dos meristemas vegetais e desenvolvem-se de acordo com as necessidades da célula, surgindo diferentes tipos de plastos como: os cromoplastos (que contêm pigmentos), os leucoplastos (sem pigmento), etioplastos (que se desenvolvem na ausência de luz), amiloplastos (que acumulam amido como substância de reserva), proteoplastos (que armazenam proteína) e os oleoplastos (acumulam lipídeos).
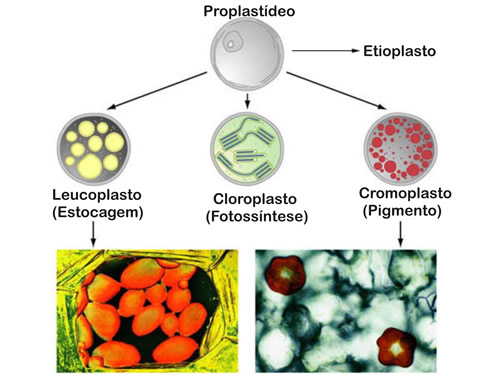
Os cloroplastos são um tipo de cromoplastos que contém pigmento chamado clorofila, que são capazes de absorver a energia eletromagnética da luz solar e a convertem em energia química por um processo chamado fotossíntese.
As células vegetais e as algas verdes possuem um grande número de cloroplastos, de forma esférica ou ovóide, variando de tamanho de acordo com o tipo celular, e são bem maiores que as mitocôndrias.
As células vegetais e as algas verdes possuem um grande número de cloroplastos, de forma esférica ou ovóide, variando de tamanho de acordo com o tipo celular, e são bem maiores que as mitocôndrias.
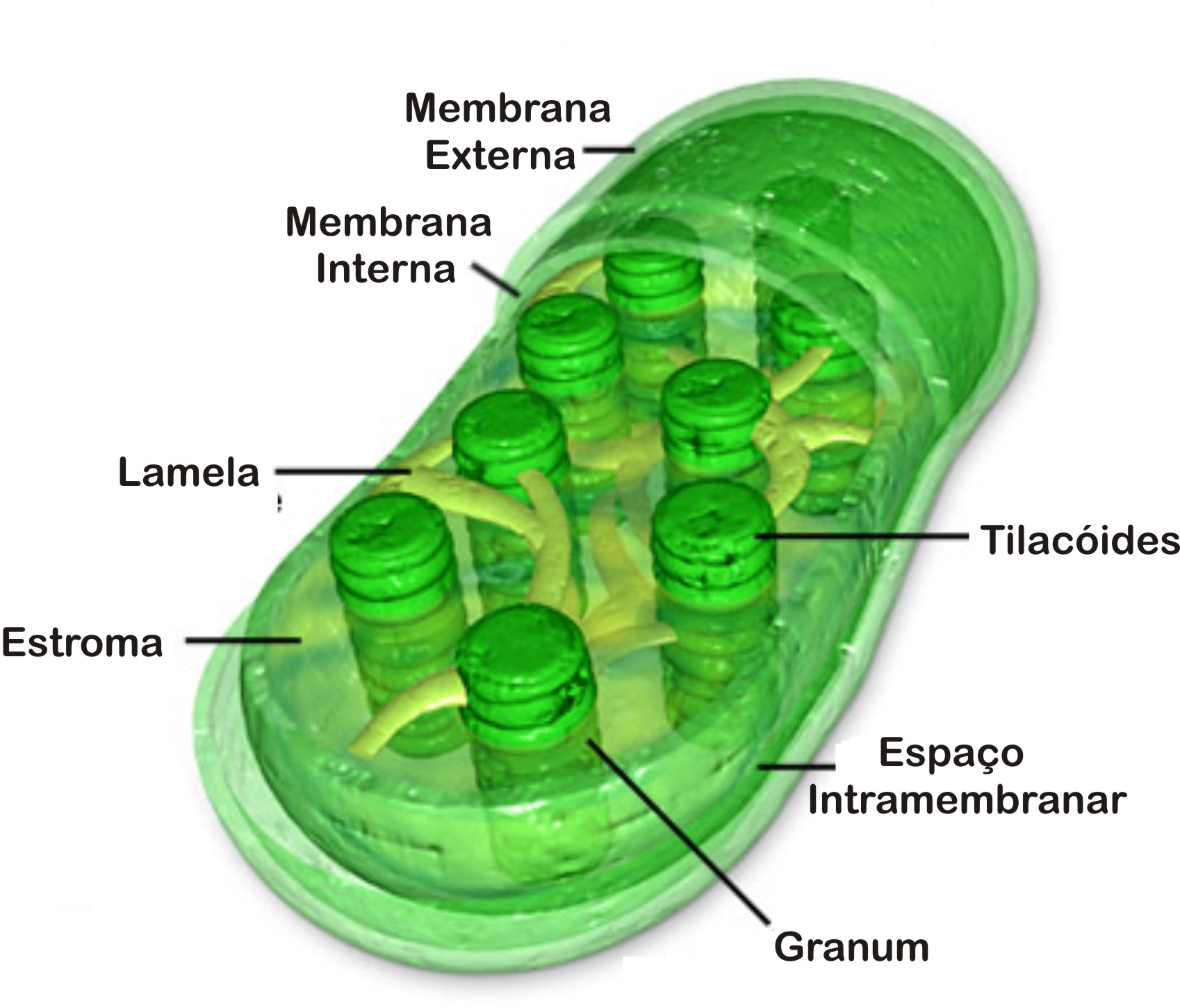
Origem evolutiva dos cloroplastos
Acredita-se que os cloroplastos tenham se originado de organismos procariontes fotossintéticos(algas azuis), que se instalaram em células primitivas eucariontes aeróbicas por endossimbiose.
Essa simbiose há cerca de 1,2 bilhões de anos, teria dado origem às algas vermelhas, depois as algas pardas e verdes e aos vegetais superiores.
Durante o processo evolutivo, as bactérias precursoras dos cloroplastos transferiram parte de seu material genético para o DNA da célula hospedeira, assim passaram a depender do genoma da célula hospedeira para a produção de muitas de suas proteínas. Esta origem é semelhante ao da mitocôndria, mas existem diferenças como o tamanho das organelas, o cloroplasto é bem maior que a mitocôndria, e a fonte de energia é diferente, o cloroplasto usa energia luminosa enquanto a mitocôndria usa energia química.
Composição química dos cloroplastos
Os cloroplastos são as organelas mais evidentes das células vegetais. Ela é composta por 50% de proteínas, 35% de lipídeos, 5% de clorofila, água e carotenóides. Parte das proteínas são sintetizadas pelo núcleo da célula, mas os lipídeos são sintetizados dentro da própria organela.
O número de cloroplastos é regulado pela célula. Existem células que contém apenas um cloroplasto, mais a maioria das células que realizam fotossíntese contém cerca de 40 a 200 cloroplastos, que se movimentam em função da intensidade de luz e da corrente citoplasmática. Semelhantes às mitocôndrias, os cloroplastos são envoltos por duas membranas, uma externa altamente permeável, e uma interna que necessita de proteínas específicas para o transporte de metabólicos, e um espaço intermembrana.
No interior da organela existe uma matriz amorfa chamada estroma que contém várias enzimas, grãos de amido, ribossomos e DNA.
| 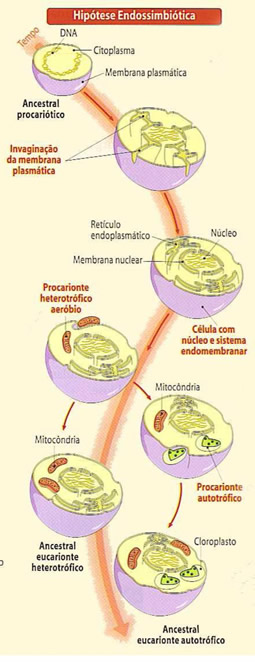 |
No entanto, a membrana interna do cloroplasto não é dobrada em cristas e não contém uma cadeia transportadora de elétrons. Mergulhado no estroma, existe um sistema de membrana (bicamada) que forma um conjunto de sacos achatados em forma de discos chamados de membrana tilacóide (do grego thylakos, saco).
O conjunto de discos empilhados recebe o nome de granum. O lúmen da membrana tilacóide é chamado de espaço tilacóide. Na membrana exposta ao estroma se localizam as clorofilas que participam da fotossíntese.
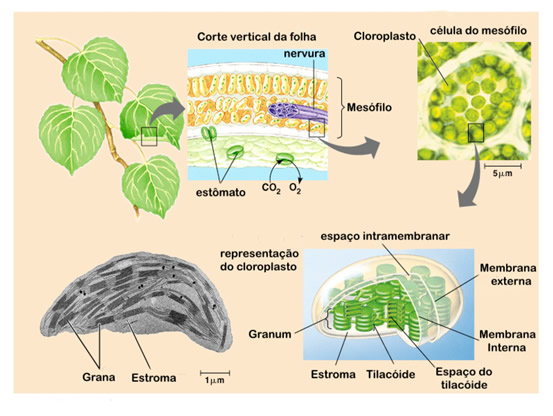
Os pigmentos ligados a diferentes proteínas e lipídeos nas membranas dos tilacóides granares e estromáticos formam sistemas complexos de proteínas-clorofila denominados fotossistemas. Há dois tipos de fotossistemas:
Fotossistema I: localizado na região da membrana voltada para o estroma, são as menores partículas intramembranosas.
Fotossistema II: localizado em tilacóides granares, formado por partículas maiores.
Fotossistema I: localizado na região da membrana voltada para o estroma, são as menores partículas intramembranosas.
Fotossistema II: localizado em tilacóides granares, formado por partículas maiores.
Sistema genético dos cloroplastos
O genoma plastidial consiste em uma pequena molécula de DNA circular, com características muito semelhantes com das mitocôndrias e das bactérias.
O DNA dos plastos ocorre em maior quantidade e é mais complexo do que da mitocôndria. Existem 30 a 200 cópias de DNA por organela contendo aproximadamente 120 genes.
O sequenciamento genético dos cloroplastos de várias plantas levou a identificação de muitos desses genes. Eles transcrevem todos os RNAs ribossômicos que compõem os plastoribossomos e 30 tipos diferentes de RNA transportadores.
Esse genoma codifica ainda 20 proteínas ribossômicas, 30 proteínas que funcionam na fotossíntese e algumas subunidades de RNA polimerase (proteínas envolvidas na expressão gênica).
O DNA dos plastos ocorre em maior quantidade e é mais complexo do que da mitocôndria. Existem 30 a 200 cópias de DNA por organela contendo aproximadamente 120 genes.
O sequenciamento genético dos cloroplastos de várias plantas levou a identificação de muitos desses genes. Eles transcrevem todos os RNAs ribossômicos que compõem os plastoribossomos e 30 tipos diferentes de RNA transportadores.
Esse genoma codifica ainda 20 proteínas ribossômicas, 30 proteínas que funcionam na fotossíntese e algumas subunidades de RNA polimerase (proteínas envolvidas na expressão gênica).
Mas mesmo sintetizando suas próprias proteínas, cerca de 90% das proteínas dos cloroplastos são codificadas pelos genes nucleares que são importadas do citosol para a organela.
Luz - Componente indispensável da fotossíntese
A luz que banha a Terra é componente do amplo espectro de radiações eletromagnéticas provenientes do Sol, e que se propagam como ondas. O modo como essas ondas se propagam depende da energia: quanto mais energia uma onda tiver, menor será seu comprimento.
Dentro do amplo espectro de radiações eletromagnéticas, apenas uma pequena parte é visível aos nossos olhos – são as radiações cujos comprimentos de onda vão de 380 a 760 nanômetros. Essas estreitas faixas de comprimento de onda da luz visível corresponde às diferentes cores que são observadas quando se faz passar a luz por um prisma, o que provoca a dispersão (separação) dessas diferentes radiações.
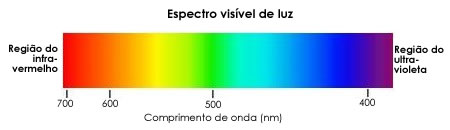
Outra característica importante da luz é a sua natureza corpuscular, ou seja, a luz é característica por incidir na forma de corpúsculos, conhecidos como fótons. Os fótons são considerados “pacotes” de energia associados a cada comprimento de onda partícula. Luz de pequeno comprimento de onda, como a luz violeta, possui fótons altamente energéticos. Luz de grande comprimento de onda, como a vermelha e a laranja, possuem fótons pouco energéticos. Assim, cada radiação luminosa, cada comprimento de onda luminosa, é portadora de uma certa energia. E o fato notável é que as plantas aproveitam essa energia para a produção de matéria orgânica na fotossíntese.
Como isso pode ser provado?
Se um feixe de luz branca passar por um prisma se decompondo em diversas cores, e em cada cor colocarmos um vidro cheio de água, lacrado, e com uma plantinha dentro, veremos com o decorrer do tempo que as plantas submetidas às radiações vermelha e azul mostram o mais alto grau de atividade fotossintética. Isso pode ser percebido pelo tamanho da bolha de oxigênio que se formou em cada vidro.

Pigmentos fotossintetisantes
O termo "pigmento" significa substância colorida. A cor do pigmento fotossintetizante depende das faixas do espectro da luz visível que ele absorve ou reflete.
A clorofila, que dá a cor verde característica da maioria dos vegetais, absorve muito bem a luz nas faixas do vermelho e do violeta, refletindo a luz verde.
Como a luz refletida é a que atinge os nossos olhos, essa é a cor que vemos, ao olharmos para uma folha. O perfil de absorção de luz de uma substância é o seu espectro de absorção.
Todas as células fotossintetizantes, exceto as bacterianas, contêm 2 tipos de clorofila, e um deles sempre é a clorofila a. O segundo tipo de clorofila geralmente é a clorofila b (nos vegetais superiores) ou a clorofila c (em muitas algas). Esses diversos tipos de clorofila diferem quanto à faixa do espectro da luz visível na qual cada uma delas capta luz com mais eficiência.
Todas as células fotossintetizantes, exceto as bacterianas, contêm 2 tipos de clorofila, e um deles sempre é a clorofila a. O segundo tipo de clorofila geralmente é a clorofila b (nos vegetais superiores) ou a clorofila c (em muitas algas). Esses diversos tipos de clorofila diferem quanto à faixa do espectro da luz visível na qual cada uma delas capta luz com mais eficiência.
As clorofilas a e b possuem espectros de absorção de luz ligeiramente diferentes, como mostra o gráfico a seguir:
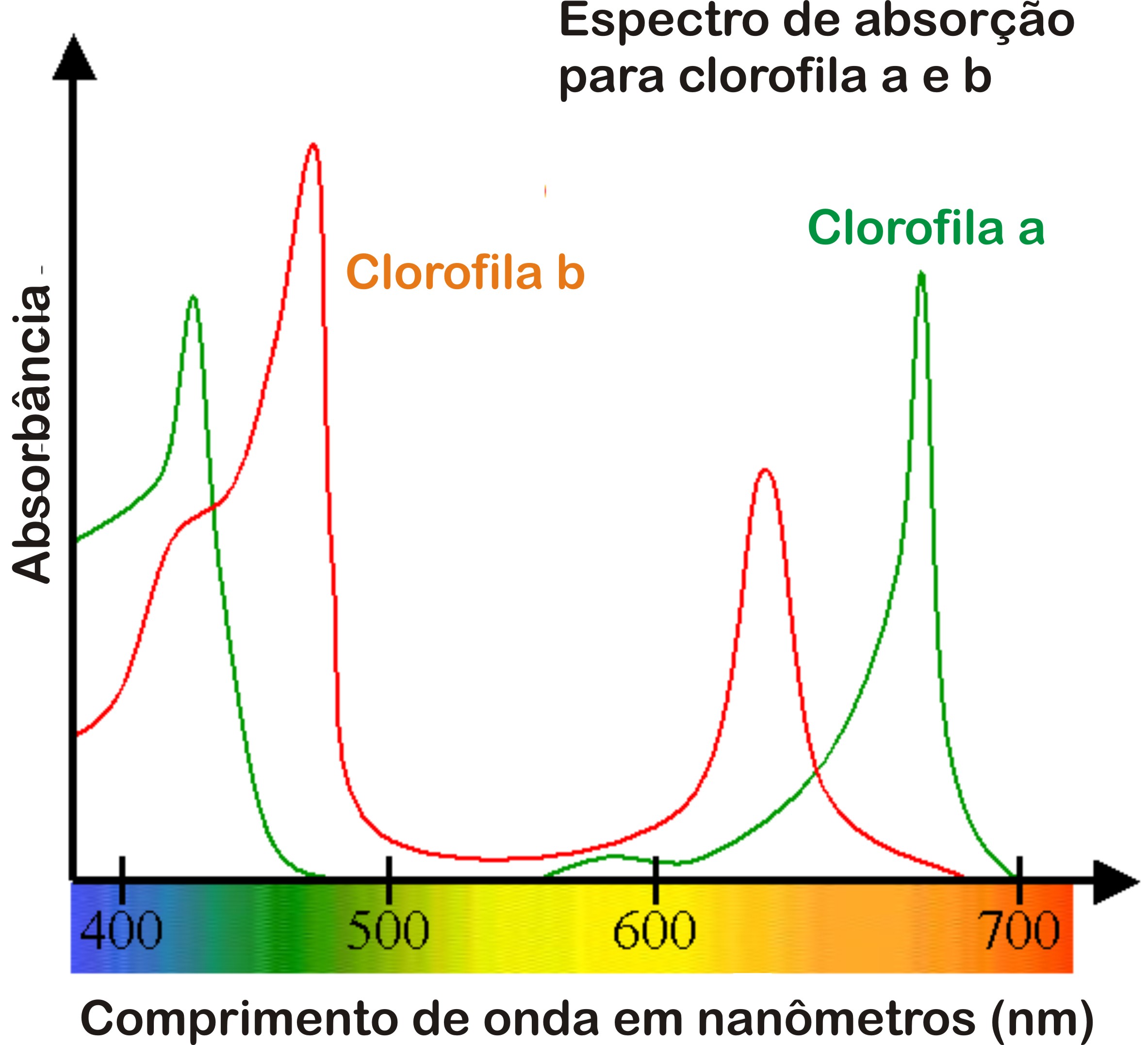
Podemos verificar, analisando o gráfico, que ambas as clorofilas possuem dois picos de absorção: um mais elevado, na faixa do violeta, e um outro menor, na faixa do vermelho.
Os carotenóides são pigmentos acessórios. Eles absorvem luz em faixas um pouco diferentes das faixas das clorofilas. A presença desses pigmentos acessórios faz com que muitas folhas tenham cores diferentes do verde. Embora tenham clorofila, a presença desses outros pigmentos em grandes quantidades mascara a sua presença e deixa as folhas com outras cores (arroxeadas, alaranjadas, amarelas, etc.).
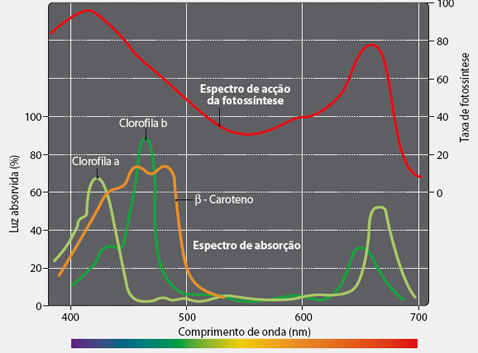
Muitas folhas mudam de cor, no inverno, pela diminuição na quantidade de clorofila. Como a quantidade dos outros pigmentos não se altera tão significativamente, as suas cores passam a ser vistas, tornando as folhas geralmente amareladas.
As etapas da fotossíntese
A fotossíntese ocorre em duas grandes etapas, que envolvem várias reações químicas: a primeira é afase clara (também chamada de fotoquímica) e a segunda é a fase escura (também conhecida como fase química).
Em linhas gerais, os eventos principais da fotossíntese são a absorção da energia da luz pela clorofila; a redução de uma aceptor de elétrons chamado NADP, que passa a NADPH2; a formação de ATP e asíntese de glicose.
Em linhas gerais, os eventos principais da fotossíntese são a absorção da energia da luz pela clorofila; a redução de uma aceptor de elétrons chamado NADP, que passa a NADPH2; a formação de ATP e asíntese de glicose.
A fase escura da fotossíntese não precisa ocorrer no escuro. O que o nome quer indicar é que ela ocorre mesmo na ausência de luz – ela só precisa de ATP e NADH2 para ocorrer.
Fase clara ou fotoquímica: Quebra da água e liberação de oxigênio
Esta fase ocorre na membrana dos tilacóides e dela participam um complexo de pigmentos existente nosgrana, aceptores de elétrons, moléculas de água e a luz. Como resultado desta fase temos a produção deoxigênio, ATP (a partir de ADP + Pi) e também a formação de uma substância chamada NADPH2;.Tanto o ATP quanto o NADPH2; serão utilizadas na fase escura.
Na fase clara, a luz penetra nos cloroplastos e atinge o complexo de pigmentos, ao mesmo tempo em que provoca alterações nas moléculas de água. De que maneira essa ação da luz resulta em produtos que podem ser utilizadas na segunda fase da fotossíntese?
Um dos acontecimentos marcantes da fase clara são as chamadas fotofosforilações cíclica e acíclica.
Na fotofosforilação cíclica, ao ser atingida pela luz do Sol, a molécula de clorofila libera elétrons. Esses elétrons são recolhidos por determinadas moléculas orgânicas chamadas aceptores de elétrons, que os enviam a uma cadeia de citocromos (substâncias associadas ao sistema fotossintetizante e que são assim chamadas por possuírem cor). Daí, os elétrons retornam à clorofila.
Na fotofosforilação cíclica, ao ser atingida pela luz do Sol, a molécula de clorofila libera elétrons. Esses elétrons são recolhidos por determinadas moléculas orgânicas chamadas aceptores de elétrons, que os enviam a uma cadeia de citocromos (substâncias associadas ao sistema fotossintetizante e que são assim chamadas por possuírem cor). Daí, os elétrons retornam à clorofila.
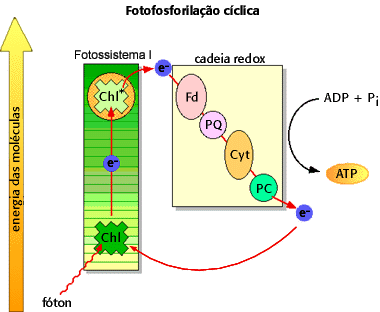
A resposta é que ao efetuar o retorno para a molécula de clorofila, a partir dos citocromos, os elétrons liberam energia, pois retornam aos seus níveis energéticos originais. E essa energia é aproveitada para a síntese de moléculas de ATP, que serão utilizadas na fase escura da fotossíntese.
Perceba que o caminho executado pelos elétrons é cíclico. Por esse motivo, costuma-se denominar essa via de fotofosforilação cíclica, devido à ocorrência de síntese de inúmeras moléculas de ATP em um processo cíclico, com a participação da luz e de moléculas de clorofila.
Ao mesmo tempo que isso ocorre, moléculas de água – ao serem atingidas pela luz do Sol – são “quebradas” (usa-se o termo “fotólise da água” para designar a quebra das moléculas de água) eliberam prótons (H+), elétrons (e-) e moléculas de oxigênio. Os prótons são captados por moléculas de NADP, que se convertem em NADPH2; moléculas de oxigênio são liberados para o meio; e os elétrons voltam para a clorofila, repondo aqueles que ela perdeu no início do processo.
Perceba que o caminho executado pelos elétrons é cíclico. Por esse motivo, costuma-se denominar essa via de fotofosforilação cíclica, devido à ocorrência de síntese de inúmeras moléculas de ATP em um processo cíclico, com a participação da luz e de moléculas de clorofila.
Ao mesmo tempo que isso ocorre, moléculas de água – ao serem atingidas pela luz do Sol – são “quebradas” (usa-se o termo “fotólise da água” para designar a quebra das moléculas de água) eliberam prótons (H+), elétrons (e-) e moléculas de oxigênio. Os prótons são captados por moléculas de NADP, que se convertem em NADPH2; moléculas de oxigênio são liberados para o meio; e os elétrons voltam para a clorofila, repondo aqueles que ela perdeu no início do processo.
A Etapa Fotoquímica da Fotossíntese
Também é chamada "fase clara" da fotossíntese, uma vez que a sua ocorrência é totalmente dependente da luz. Como se trata de uma etapa que conta com a participação das moléculas de clorofila, acontece no interior dos tilacóides, em cujas faces internas de suas membranas as moléculas desse pigmento fotossintetizante estão "ancoradas".
Nessa etapa, a clorofila, ao ser iluminada, perde elétrons, o que origina "vazios" na molécula. O destino dos elétrons perdidos e a reocupação desses vazios podem obedecer a 2 mecanismos distintos, chamadosfotofosforilação cíclica e fotofosforilação acíclica.
A - Fotofosforilação cíclica
No chamado fotossistema I, predomina a clorofila a. Essa, ao ser iluminada, perde um par de elétrons excitados (ricos em energia). Estabelece-se, na molécula da clorofila, um "vazio" de elétrons. O par de elétrons é recolhido por uma série de citocromos, substâncias que aceitam elétrons adicionais, tornando-se instáveis e transferindo esses elétrons para outras moléculas.
À medida que passam pela cadeia de citocromos, os elétrons vão gradativamente perdendo energia, que é empregada na fosforilação (produção de ATP pela união de mais um grupo de fosfato a uma molécula de ADP). Como essa fosforilação é possível graças à energia luminosa, captada pelos elétrons da clorofila, é chamada fotofosforilação.
À medida que passam pela cadeia de citocromos, os elétrons vão gradativamente perdendo energia, que é empregada na fosforilação (produção de ATP pela união de mais um grupo de fosfato a uma molécula de ADP). Como essa fosforilação é possível graças à energia luminosa, captada pelos elétrons da clorofila, é chamada fotofosforilação.
Após a passagem pela cadeia de citocromos, os elétrons retornam à molécula da clorofila, ocupando o "vazio" que haviam deixado. Como os elétrons retornam para a clorofila, o processo é cíclico.
B - Fotofosforilação acíclica
Esse mecanismo emprega dois sistemas fotossintetizantes: o fotossistema I e o fotossistema II. No fotossistema I, predomina a clorofila a, enquanto no fotossistema II, predomina a clorofila b.
A clorofila a, iluminada, perde um par de elétrons ativados, recolhidos por um aceptor especial, aferridoxina. Ao mesmo tempo, a clorofila b, excitada pela luz, perde um par de elétrons que, depois de atravessarem uma cadeia de citrocromos, ocupa o "vazio" deixado na molécula da clorofila a. Durante a passagem desses elétrons pela cadeia de citocromos, há liberação de energia e produção de ATP (fosforilação). Como o "vazio de elétrons" da clorofila a não é preenchido pelos mesmos elétrons que saíram dessa molécula, o mecanismo é chamado fotofosforilação acíclica.
Esse mecanismo emprega dois sistemas fotossintetizantes: o fotossistema I e o fotossistema II. No fotossistema I, predomina a clorofila a, enquanto no fotossistema II, predomina a clorofila b.
A clorofila a, iluminada, perde um par de elétrons ativados, recolhidos por um aceptor especial, aferridoxina. Ao mesmo tempo, a clorofila b, excitada pela luz, perde um par de elétrons que, depois de atravessarem uma cadeia de citrocromos, ocupa o "vazio" deixado na molécula da clorofila a. Durante a passagem desses elétrons pela cadeia de citocromos, há liberação de energia e produção de ATP (fosforilação). Como o "vazio de elétrons" da clorofila a não é preenchido pelos mesmos elétrons que saíram dessa molécula, o mecanismo é chamado fotofosforilação acíclica.
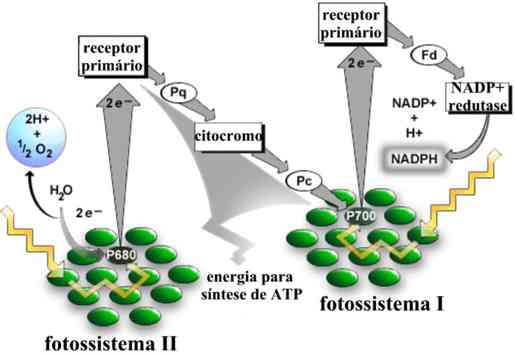
No interior dos cloroplastos, a água é decomposta na presença da luz. Essa reação é a fotólise da água.(ou reação de Hill).
Dos produtos da fotólise da água, os elétrons vão ocupar os "vazios" deixados pela perda de elétrons pela clorofila b. Os prótons H+, juntamente com os elétrons perdidos pela clorofila a, irão transformar o NADP (nicotinamida-adenina-dinucleotídeo fosfato) em NADPH. Ao mesmo tempo, oxigênio é liberado. Esse é um aspecto importante da fotossíntese: todo o oxigênio gerado no processo provém da fotólise da água.
Os seres fotossintetizantes utilizam a água como fonte de átomos de hidrogênio para a redução do NADP. Esses átomos de hidrogênio são posteriormente empregados na redução do CO2 até carboidrato. A equação geral do processo é a seguinte:
O valor n corresponde, geralmente, a seis, o que leva à formação de glicose (C6H12O6). Entretanto, como todo oxigênio liberado vem da água, a equação deve ser corrigida para:
Dessa forma, pode-se explicar a origem de uma quantidade 2n de átomos de oxigênio a partir de uma quantidade de 2n moléculas de água (H2O).
Fase escura ou química: Produção de Glicose
Nessa fase, a energia contida nos ATP e os hidrogênios dos NADPH2, serão utilizados para a construção de moléculas de glicose. A síntese de glicose ocorre durante um complexo ciclo de reações (chamado ciclo das pentoses ou ciclo de Calvin-Benson), do qual participam vários compostos simples.
Durante o ciclo, moléculas de CO2 unem-se umas as outras formando cadeias carbônicas que levam à produção de glicose. A energia necessária para o estabelecimento das ligações químicas ricas em energia é proveniente do ATP e os hidrogênio que promoverão a redução dos CO2 são fornecidos pelos NADPH2.
Veja com mais detalhes o ciclo de Calvin
O Ciclo de Calvin
O ciclo começa com a reação de uma molécula de CO2 com um açúcar de cinco carbonos conhecido comoribulose difosfato catalisada pela enzima rubisco (ribulose bifosfato carboxilase/oxigenase, RuBP), uma das mais abundantes proteínas presentes no reino vegetal.
Forma-se, então, um composto instável de seis carbonos, que logo se quebra em duas moléculas de três carbonos (2 moléculas de ácido 3-fosfoglicérico ou 3-fosfoglicerato, conhecidas como PGA). O ciclo prossegue até que no final, é produzida uma molécula de glicose e é regenerada a molécula de ribulose difosfato.
Forma-se, então, um composto instável de seis carbonos, que logo se quebra em duas moléculas de três carbonos (2 moléculas de ácido 3-fosfoglicérico ou 3-fosfoglicerato, conhecidas como PGA). O ciclo prossegue até que no final, é produzida uma molécula de glicose e é regenerada a molécula de ribulose difosfato.
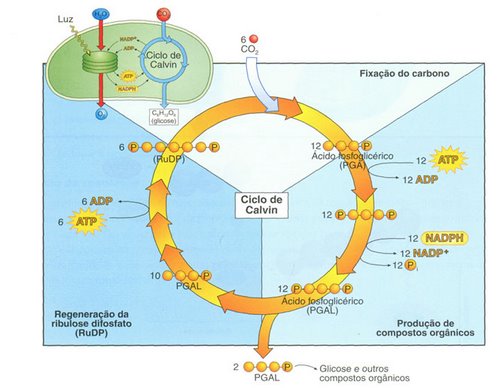
Note, porém, que para o ciclo ter sentido lógico, é preciso admitir a reação de seis moléculas de CO2 com seis moléculas de ribulose difosfato, resultando em uma molécula de glicose e a regeneração de outras seis moléculas de ribulose difosfato.
A redução do CO2 é feita a partir do fornecimento de hidrogênios pelo NADH2 e a energia é fornecida pelo ATP. Lembre-se que essas duas substâncias foram produzidas na fase clara.
O esquema apresentado é uma simplificação do ciclo de Clavin: na verdade, as reações desse ciclo se parecem com as que ocorrem na glicólise, só que em sentido inverso.
É correto admitir, também, que o ciclo origina unidades do tipo CH2O, que poderão ser canalizadas para a síntese de glicose, sacarose, amido e, inclusive, aminoácidos, ácidos graxos e glicerol.
Fatores que influenciam a fotossíntese
A intensidade com a qual uma célula executa a fotossíntese pode ser avaliada pela quantidade de oxigênio que ela libera para o ambiente, ou pela quantidade de CO2 que ela consome.
Quando se mede a taxa de fotossíntese de uma planta, percebe-se que essa taxa pode aumentar ou diminuir, em função de certos parâmetros. Esses parâmetros são conhecidos como fatores limitantes da fotossíntese. A fotossíntese tem alguns fatores limitantes, alguns intrínsecos e outros extrínsecos.
Fatores limitantes intrínsecos
Disponibilidade de pigmentos fotossintetizantes
Como a clorofila é a responsável principal pela captação da energia limunosa, a sua falta restringe a capacidade de captação da energia e a possibilidade de produzir matéria orgânica.
Disponibilidade de enzimas e de cofatores
Todas as reações fotossintéticas envolvem a participação de enzimas e de co-fatores, como os aceptores de elétrons e os citocromos. A sua quantidade deve ser ideal, para que a fotossíntese aconteça com a sua intensidade máxima.
Todas as reações fotossintéticas envolvem a participação de enzimas e de co-fatores, como os aceptores de elétrons e os citocromos. A sua quantidade deve ser ideal, para que a fotossíntese aconteça com a sua intensidade máxima.
Fatores limitantes extrínsecos
A concentração de CO2
O CO2 (gás carbônico ou dióxido de carbono) é o substrato empregado na etapa química como fonte do carbono que é incorporado em moléculas orgânicas. As plantas contam, naturalmente, com duas fontes principais de CO2: o gás proveniente da atmosfera, que penetra nas folhas através de pequenas aberturas chamadas estômatos, e o gás liberado na respiração celular.
Sem o CO2, a intensidade da fotossíntese é nula. Aumentando-se a concentração de CO2 a intensidade do processo também se eleva. Entretanto, essa elevação não é constante e ilimitada. Quando todo o sistema enzimático envolvido na captação do carbono estiver saturado, novos aumentos na concentração de CO2 não serão acompanhados por elevação na taxa fotossintética.
|
A Temperatura
Na etapa química, todas as reações são catalisadas por enzimas, e essas têm a sua atividade influenciada pela temperatura.
De modo geral, a elevação de 10 °C na temperatura duplica a velocidade das reações químicas.
Entretanto, a partir de temperaturas próximas a 40 °C, começa a ocorrer desnaturação enzimática, e a velocidade das reações tende a diminuir. Portanto, existe uma temperatura ótima na qual a atividade fotossintetizante é máxima, que não é a mesma para todos os vegetais. |
O comprimento de onda
A assimilação da luz pelas clorofilas a e b, principalmente, e secundariamente pelos pigmentos acessórios, como os carotenóides, determina oespectro de ação da fotossíntese.
Nota-se a excelente atividade fotossintética nas faixas do espectro correspondentes à luz violeta/azul e à luz vermelha, e à pouca atividade na faixa do verde. Para que uma planta verde execute a fotossíntese com boa intensidade, não se deve iluminá-la com luz verde, uma vez que essa luz é quase completamente refletida pelas folhas. |
Intensidade luminosa
Quando uma planta é colocada em completa obscuridade, ela não realiza fotossíntese. Aumentando-se a intensidade luminosa, a taxa da fotossíntese também aumenta. Todavia, a partir de um certo ponto, novos aumentos na intensidade de iluminação não são acompanhados por elevação na taxa da fotossíntese. A intensidade luminosa deixa de ser um fator limitante da fotossíntese quando todos os sistemas de pigmentos já estiverem sendo excitados e a planta não tem como captar essa quantidade adicional de luz. Atingiu-se o ponto de saturação luminosa.
Aumentando-se ainda mais a intensidade de exposição à luz, chega-se a um ponto a partir do qual a atividade fotossintética passa a ser inibida. Trata-se do ponto de inibição da fotossíntese pelo excesso de luz. |
Nenhum comentário:
Postar um comentário